マクロな生命現象を
常識にとらわれずに理解する
生物のさまざまな適応現象は驚くほど見事なものです。それゆえ、ダーウィンが進化論を提唱して以降、生物の適応進化というプロセスは、生態学者や進化生物学者をつねに魅了してきました。進化の原動力やプロセス、遺伝的基盤を明らかにすることは、生物進化の実態を解き明かすという点で生物学の発展に大きく貢献してきたことは疑う余地もありません。一方で、昨今の進化生態学や進化生物学はさまざまな進化過程やその遺伝的基盤の記載をするようなケーススターディーを積み重ねているに過ぎなくなりつつあるという現実もあるように思います(もちろん、いまなお、そのような積み重ねの中で大きな発見が続いていますが)。
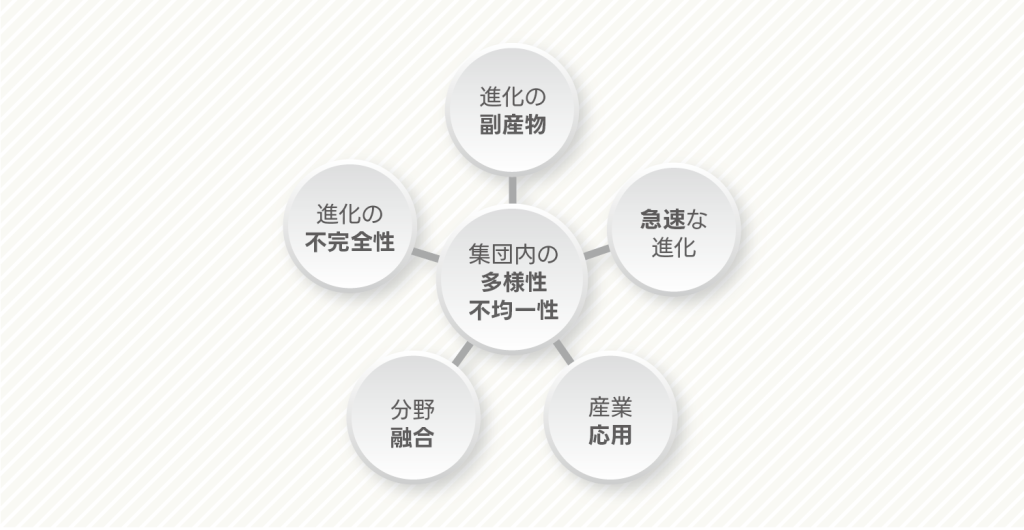
私は、進化生態学や進化生物学の停滞や閉塞感といえるようなものを打破すべく、主に「5つの視点」で研究を進めています。一つ目は、「生物が必ず最適値に向かって進化する」という暗黙の仮定や「生物が現時点でほぼ完全に環境への適応が完了していること」という仮定を切り崩すことです。すなわち、(アイデアとしては決して新しいものではありませんが)生物の適応進化が完璧ではないという考えに立脚し、「適応や多様化といった進化が制限される原因」を探ることです。2つ目は、生物進化の副次的効果や副作用を明らかにすることです。表現型の進化や遺伝的多様性の進化が群れ行動や個体群動態、群集動態、生態系機能などのよりマクロな生命現象にどのようなインパクトを与えるかを考えることです。急速な表現型進化や多様性の進化に着目しながら、進化動態と人口学的動態をリンクさせ、進化生態学や進化生物学の新たな平地を切り拓こうと考えています。3つ目は、〝自然〟が生み出す自然選択にこだわらない進化生物学を展開することです。特に急速な環境変化としての「都市化」に着目し、〝不自然選択〟が起こす生物の進化を調べることです。4つ目は、他分野と融合しながら学際的に生態学や進化学的な研究を推進することです。ゲノム情報やトランスクリプトーム情報を活用した分子生物学的なアプローチはもちろん、機械学習(逆強化学習)や流体力学などの工学的手法や、心理学な視点を進化生態学に取り入れることを通じて、今までにないアイデアや理論を提唱することを目指しています。5つ目は、進化生態学の理論を農産業や食品工業、環境問題への応用です。進化生物学や生態学の基礎研究の成果を作物生産、食品生産に応用することで、昨今の食糧問題やエネルギー問題の解決に結びつけようと考えています。

また、多様な手法、多様な材料で研究を展開しています。ゲノム解析やトランスクリプトーム解析、分子実験、フィールドワーク、行動観察、形態測定、ビッグデータ解析、シミュレーションなどのさまざまな手法を駆使しながら、動物、植物、微生物など多くの生物を使うことで、自然界に潜む生態学的あるいは進化学的に重要なさまざまな原理や一般則を探求しています。
キーワード 遺伝的多様性、個性、表現型可塑性、ゆらぎ、遺伝子流動、分布域、個体群動態、多様性の維持機構、多様性の機能、ゲノム解析、生態-進化フィードバック、個体間相互作用、環境適応、適応阻害、概潮汐リズム、都市ストレス、都市適応、逆強化学習、季節適応、急速な進化、表現型可塑性、進化の方向性、キイロショウジョウバエ、野生のショウジョウバエ、トンボ、チリメンカワニナ、ネジバナ、ウキクサ、ビッグデータ
多様性による集団の機能創発とその応用
多様性、不均一性、ヘテロ性は、生命現象を象徴する特徴です。組織や腫瘍の中の細胞には不均一性がありますし、当然、個体は多様な種類の細胞で構成されています。群れや集団は多様な個性をもった個体によって構成されていて、生態系は多様な種によって構成されています。生命システムは、どんな階層においても、多様性な要素の集合体であるわけです。このような多様な要素は非相加的にあるいは相乗的にシステム全体の特徴や機能、パフォーマンスを形作る可能性があります。私は、多様な要素間の相互作用が織りなすシステムのダイナミクス、すなわち、創発的なダイナミクスを解明することこそが生物学の醍醐味かもしれません。われわれは、多様性や不均一性に着目し、以下のようなテーマの研究を推進しています。
ショウジョウバエにおける遺伝的多様性の生態的機能とそのメタ遺伝基盤

多様性(ダイバーシティー)は、社会や集団の「力」になり、イノベーション生み出すことがヒトにおいて知られています。ヒト以外の生物でもそのようなことが起こるのでしょうか?また、多様性はいつでも集団にとっていい効果をもたらすのでしょうか?どんな多様性も集団に対して何らかの相乗的効果を生み出すのでしょうか?このような背景から、私たちは、ショウジョウバエにおける遺伝的な種内多様性が個体群動態に与える影響を調べています。ゲノム配列が既知の多数の系統(DGRP系統)を網羅的に組み合わせて飼育を行ない、組み合わせごとに個体群特性(集団の生存率やバイオマスなど)を計測することで、「多様性効果」を推定します。また、そのような効果をもたらす遺伝子の探索をゲノムワイドに探索するような研究も行なっています。同様の実験を、ショウジョウバエ属の他の種においても行なっています。
「発生ゆらぎ」による個性とその生態的機能
集団内の異質性(不均一性)を生み出す要因は、「遺伝」と「環境に誘導される表現型可塑性」であると思われがちだが、もう一つの要因に「発生ゆらぎ」があります。発生過程で生じるわずかな「ゆらぎ」の蓄積の結果として、個体間の差異が生じているのです。例えば、脳の発生のゆらぎによって生じる行動パターンの個体差が生まれたりします。一方で、ゆらぎによって生じる個体差の生態的機能はまったくわかっていません。私たちは、行動に見られる発生ゆらぎの程度を多数のショウジョウバエ系統で評価するとともに、そのようなゆらぎの生態的機能や進化機構を調べています。
遺伝的多様性がショウジョウバエの群れ行動や群れのパフォーマンスに与える影響

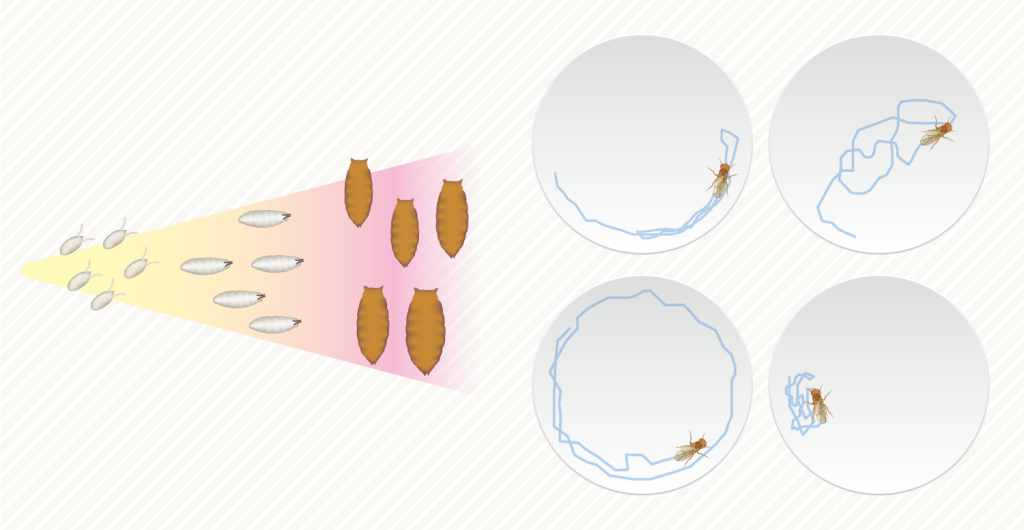
集団、あるいは個体群より小さいスケールの集合として「群れ」があります。程度の差こそあれ、多くの動物が群れをなして生活しています。群れることで捕食リスクを低減したりできるためです。一方で、群れの研究の多くは、群れ内に存在する変異を無視している、あるいはノイズとして扱っているという現状があります。でも、実際には、ほとんどの生物で、無視できないほどの多様性や個性が群れの中には存在します。そこで私たちは、多様な個性をもった個体によって群れを構成(=さまざまな構成の群れを人工的に合成)し、多様性が個体レベルの行動や群れ全体としての振る舞いや集団的知性に与える影響を調べています。個体レベルでは解決不可能な複数行動形質間のトレードオフ関係に与える影響も調べています。材料には、キイロショウジョウバエのDGRP系統やキハダショウジョウバエ、タカハシショウジョウバエなどを使っています。なお、遺伝的な個性だけでなく、環境依存的に現れる可塑的な個体差や発生のゆらぎによって生じる個体差にも着目しながら群れ内での多様性の機能を調べています。DGRP系統を用いた解析では、群れ行動のゲノム基盤も調べています。
出芽酵母にける遺伝的多様性と個体群特性の関係とその産業応用

酵母は増殖速度が非常に早く、個体群動態を観察することが容易な生物です。マイクロプレートリーダーなどを使えば、ハイスループットで個体群増殖過程を定量することができます。実は、生態学的な解析や実験に向いている生物です。一方で、酵母は発酵や醸造に用いられるほか、バイオエタノールの生産などにも使われる「有用生物」でもあります。このような生物の増殖速度を制御したり、代謝産物の生産量を制御できるようになれば、さまざまな産業に応用できる可能性があります。私たちは、糖代謝能力の種内変異株を共培養する実験や、出芽酵母のさまざまなエコタイプの株を共培養する実験を行ない、多様性による増殖能力の改変や多様性効果の出現条件を探索しています。実際、培養条件や組み合わせを変えることで、さまざまな強さや方向性の多様性効果が出現することがわかってきました。既存の系統を組わせることによって単一系統では達成できないような高い増速特性を示したり、新奇の特性を生み出したりできる可能性を示しています。人為選択や品種改良によって「個」の生産性や頑健性を高めようとしてきた既存の方法とは異なり、「集団」をデザインすることで、発酵特性を向上させられるようになるかもしれません。
ウキクサにおける遺伝的多様性の生態的機能と機能的集団デザイン

農作物のほとんどすべては、画一栽培されています。すなわち、田んぼや畑には、単一の品種のみが栽培されています。遺伝的に均一な個体間では要求する栄養も同じでしょうから、極めて強い個体間競争が起きていると想像できます。一方で、種内の遺伝的多様性を利用した生産性やその安定性の向上は、農業や畜産業などに応用できるポテンシャルをもっています。一方で、生産性や安定性を向上させるような効果は、どのような組み合わせでも現れるというわけでありません。すなわち、多様性効果をうまく引き出すような組み合わせを見つけることが重要になります。私たちはその第一歩として、実験室環境でも培養が可能で、個体群増殖のパターンを観察しやすく、かつ、利用価値の高い植物として、ウキクサ類に着目した研究を行なっています。生態的実験とゲノム解析を組み合わせたアプローチで、機能的集団をデザインする技術を探索しようしています。
遺伝的多様性ー生態的機能関係の理論的予測
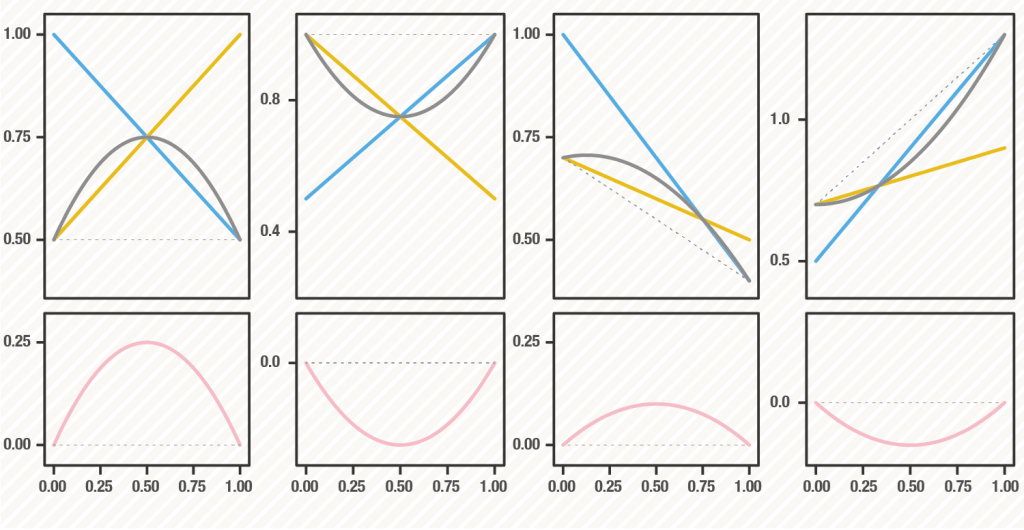
遺伝的多様性の進化(維持)と遺伝的多様性の生態的波及効果は、密接に関係しています。遺伝的多様性には、平衡選択(負の頻度依存選択やヘテロ超優性)によって恒久的に維持されるようなものもあれば、突然変異と遺伝的浮動のバランスによって中立的に残っているだけの多様性もあります。あるいは、方向性選択や多数派が有利になるような正の頻度依存選択によって近い将来消滅してしまうような一時的な多様性もあります。理論的には、多様性を維持するような平衡選択が働いている条件では多様性は相乗的に集団全体の生産性を高め(正の多様性効果)、多様性を失わせるような選択圧(正の頻度依存選択)が働いている条件では多様性は集団全体の生産性を低める(負の多様性効果)ことがわかっています。ただし、多様性を維持する原動力は、平衡選択(狭義)だけではないですし、多様性効果が生まれるメカニズムをさまざまなものがあります。どのような選択圧の働くどのような形質のどのような多様性がどのような多様性効果を生み出すのか、理論的に調べていきたいと思っています。
ビッグデータを用いた多様性の生態的効果の解析

昆虫を使った室内実験だとせいぜい数世代、酵母を使った実験でも数十世代の個体群動態しかわかりません。しかも、試験管の中で繰り広げられるダイナミクスしかわかりません。遺伝的多様性の全球レベルでの生態的なスケールアップ効果を明らかにするには、系統関係や分布域、生態的特性、形態的特性、絶滅リスク、個体群の盛衰傾向に関するビッグデータを使うことが有効です。昆虫や脊椎動物、鳥類などのさまざまな生物の全球レベルでの分布データなどを活用して、種内(集団内)多様性の進化のマクロ生物学的な波及効果を調べています。
進化の制約機構と進化の限界
生物の多様さを見ていると、生物はさまざまな環境要因に曝され、あらゆる適応進化を遂げているようにみえます。しかし、それは真実でありません。生物は遺伝子レベルや発生レベルのさまざまな「しがらみ」により、表現可能な形態や生態が制約され、結果として、進化できる方向性も強く制約されている可能性があります。また、たとえそのような制約がない場合にも、単純に集団内で遺伝的多様性が失われたり、集団外から非適応的な変異が流入してくることによって適応進化が妨げられる可能性があります。私たちは、このような「進化の制約機構」を理解することが生物進化の真の理解に重要であると考えています。そのような背景のもと、以下のような研究を展開しています。
表現型変異の制約と変異パターンの階層間での類似性と進化の予測
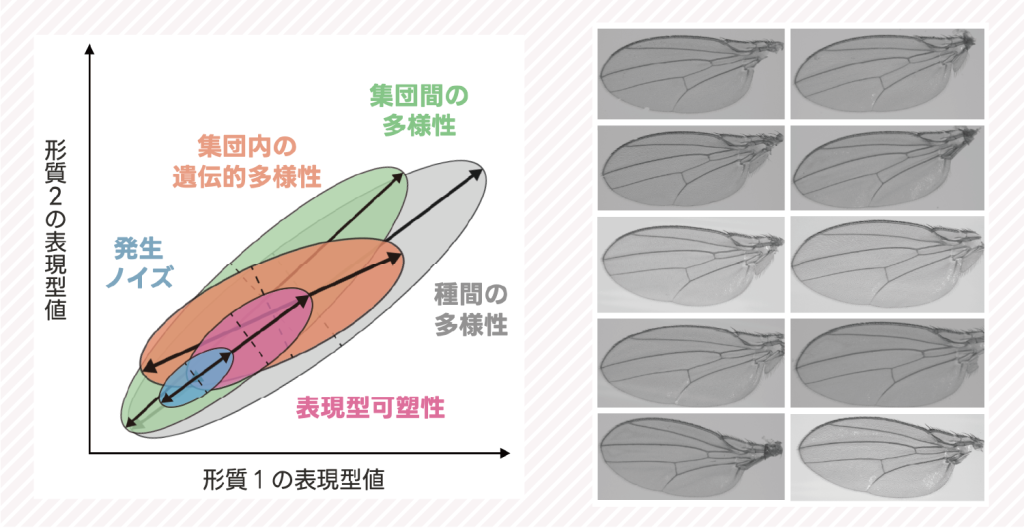
生物の形態的多様性には、階層性があります。種間での多様性がもっとも上位で、その下には、集団間での多様性があり、さらに、集団内の個体間での遺伝的多様性、同じ遺伝子型内での環境間での多様性、発生ゆらぎよって生じる多様性が続きます。このような多様性には類似性はあるのでしょうか?もし、低次の多様性が高次の多様性の成立を制約するのであれば、類似性があるいかもしれません。あるいは、いずれの階層にも共通の制約があるのであれば、階層間で類似性があってもおかしくありません。私たちは、トンボ類の翅形態、ショウジョウバエ類の翅形態、ショウジョウバエ類の遺伝子発現パターンに着目し、「発生ゆらぎ」から「大進化」までで見られる表現型変異のパターンを網羅的に比較する研究を進めています。これらを通じて、現在の低次の多様性(例えば発生ゆらぎや表現型可塑性による表現型変異)から進化や多様化が予測可能であるかどうかを調べています。
遺伝子流動の過不足と適応進化の不完全性

遺伝的多様性が枯渇するときも、遺伝的多様性が高まりすぎるときも、適応進化が制限されることが知られています。生物の分布域の辺縁(分布限界)では、遺伝子流動の相反する2つの効果によって適応進化が制限されることが知られています。
一つ目は、遺伝子流動の不足です。分布限界付近は生息に適した環境ではないので、集団は縮小したり、分断化したりします。すると、遺伝的浮動が強く働いたり、集団間での移住は制限さることで、各サブ集団内の遺伝的変異が枯渇します。私達は、アオモンイトトンボの分布北限において、遺伝的多様性の枯渇と適応進化の失敗の痕跡を検出してきました。
二つ目は、過度な遺伝子流動による最適化の阻害です。空間的な環境勾配が急峻な場合、隣接したサブ集団間でも異なった表現型が最適値となる場合があります。たとえば、河川では、標高に応じてさまざまな水環境や生物的環境に適応する必要があります。一方で、河川では、上流から下流への非対称でかつ非常に強い遺伝子流動が存在するので(洪水などを通じて)、下流側の集団では上流からの非適応的な遺伝子の流入によって遺伝的な汚染を受けることになります(=移住に伴う遺伝荷重、あるいは移住荷重)。このような荷重が環境への十分な適応を妨げる可能性がありました。私たちは、河川のさまざま環境に生息するチリメンカワニナを用いたフィールドワークと遺伝子発現解析、集団遺伝学的的解析を通じて、河川内では上流から下流への非対称な遺伝子流動が起きていることや、急峻で遺伝子流動が生じやすい河川ほど下流側での汽水環境への適応が妨げられていることを明らかにしました。 生物の分布限界や異なった環境が隣接する部分では、遺伝的多様性の不足あるいは過剰によって、さまざまな「適応進化の失敗」が存在していそうです。
急速な進化とイノベーティブな進化
生物の進化は、これまで考えられてきたよりも「速い」ことがわかってきています。「進化」は数万年とか数千年で起こる現象ではなく、数十年とか数年で起きる現象であることが明らかになりつつあります。加えて、生物は、急激な環境変化に対して劇的な進化を遂げていることがあります。私たちは、これらの問題に対して、都市化に応答した急速な進化、季節変化に対する超高速な進化、生物時計の進化に着目して以下のような研究を進めています。
都市化に伴った生物の迅速な適応と可塑的応答

近年、地球上の都市の面積は増加の一途をたどり、都市化による気温上昇(温害)や夜間人工光の増加(光害)、騒音の増大、電磁波の増加は、生物に多大なる影響を与えています。これらの急速で急激な環境変化や環境ストレスは、生物の衰退や絶滅をもたらす可能性もありますし、選択圧となって生物進化を促す可能性あります。生物の私たちは、都市に生息するショウジョウバエを用いて都市において生じた進化的な変化や可塑的な変化を検出しようとしています。具体的には、都市から郊外にかけてのさまざまな地点で採集した個体をもとに系統をたくさんつくり、それらを比較することで、都市-郊外勾配に沿った進化の痕跡の検出を試みています。また、温度や光、音、電磁波のストレスを与えたときの影響を表現型レベルや遺伝子レベルで調べています。材料は主に、オウトウショウジョウバエを使用しています。
概潮汐リズムの遺伝基盤の解明

生物は、自らが生息する環境に適応したさまざまな内在リズム(生物時計)をもっています。潮間帯や汽水域では、12.4時間周期で水深や塩分が変動するため、このような環境に生息する生物は概潮汐リズムという約12.4時間の活動リズムをもつことが知られています。しかし、概潮汐リズムという「12.4時間周期で時を刻むこと」の遺伝的基盤はまったくわかっていません。私たちは、チリメンカワニナにおいて汽水域の集団でのみ活動リズムに概潮汐リズムが存在することを発見しました。本種やその近縁種はいずれも淡水性の生物ですので、これらの事実は、本種において、比較的最近、新しい内在リズムが獲得されたことを意味します。この現象に着目し、トランスクリプトーム解析や行動解析から概潮汐リズムの獲得の進化的機構の解明を目指しています。本研究は横溝 匠さんとの共同研究として行なっています。
形質の季節間での迅速な進化

生物は、変わりゆく季節、すなわち、急速に変化する選択圧のもとで生存しています。年に1化生の昆虫や、数年に渡って生存する生物ではあまり関係ありませんが、年に何度も世代を回す生物では、世代間で受ける選択圧が劇的に変化することになります。このような選択圧の季節変化は、形質の季節変化にどれほど影響を与えるかは十分にわかっていません。私たちは、ショウジョウバエ類を材料に集団内の遺伝的な形質変異やその変異の季節間での入れ替わりがどれほど起きているのかを検証することで、迅速な進化の検証を行なうとともに、集団内の変異が変動環境下でどのような機能を果たしているかを明らかにすることを目指して研究をしています。
ネジバナの花序形態の多様性と錯視

多様性の成立メカニズムとして有名なのが、遺伝的変異と表現型可塑性です。これらの2つが注目されがちですが、もう一つの出現メカニズムがあります。それが、発生のゆらぎです。遺伝的背景も、発育環境もまったく同じだとしても、個体発生の過程で各形質の表現型はわずかに揺らぐのです。ネジバナでは、発生のゆらぎに起因する花序形態の明瞭な多型が存在します。発生ゆらぎによって生じる多様性にも、遺伝的変異と同様に副産物的な効果があるのでしょうか?私たちは、ネジバナの詳細な形態観察と、生態的調査を通じて、発生ゆらぎの生態的機能の解明を目指しています。
異分野の視点と手法の融合
ゲノムシーケンスの技術やイメージングの技術など、生物学で使われる手法も日進月歩で、生物学では新たな発見が日々続いています。一方、私たちは、生物学ではあまり使われない手法やアイデアを取り入れることを大切にしています。そこにはブレークスルーがあるような気がするからです。
逆強化学習を用いた生物の集団行動の解析
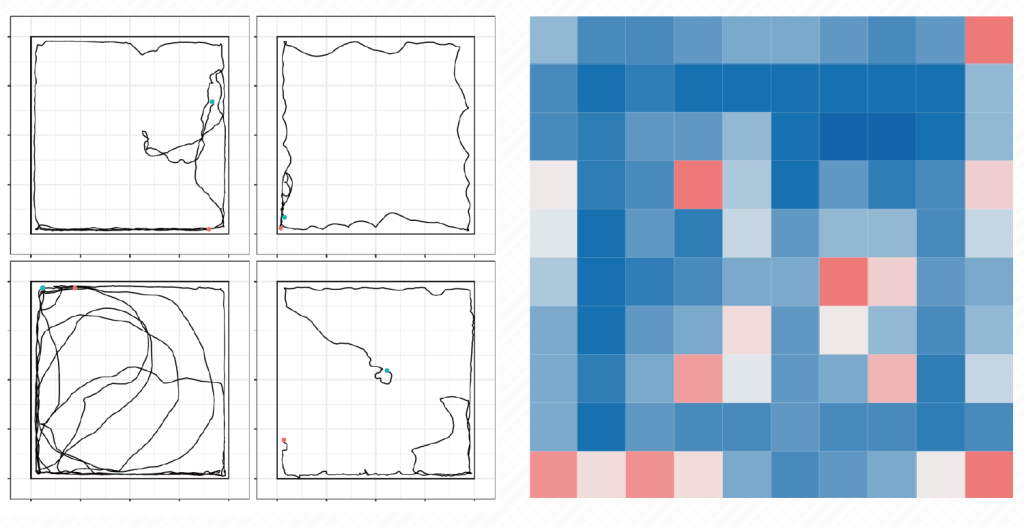
動物の個体数の動態は、一般に、湿度や温度などの無機的環境の影響(無機的環境)と、同種他個体やギルド内の異種個体、天敵、餌種との相互作用(生物的環境)によって形作られます。ですので、個体群動態や群集動態を考えるうえで、動物がどのように振る舞い、どのように他個体と相互作用するのかを理解することが実は重要になります。動物の振る舞いが正確にわかれば、ボトムアップ的に生物の個体群の動態が理解できるかもしれません。とはいえ、生物学における動物の研究のほとんどは、人による直接的な観察で動物の行動を記載してきたため、行動や移動の背景にある複雑な意思決定の過程を正確に推定・評価することは不可能でした。一方で、近年、機械学習に関わる技術的革新により、動物の移動パターンを定量・分類することが可能になってきた。とくに、逆強化学習を用いた解析により、移動軌跡のデータから、その意思決定に関わる要因(報酬)の推定や行動の予測が可能になってきている。そこで、わたしたちは、逆強化学習を用いることで、生物の行動・移動に関わる意思決定は状況依存的どのように変化するのかを明らかにしようとしています。材料として、ミールワームやショウジョウバエを用いています。
トンボ類における翅形態の進化の幾何学的解析

トンボ類は現存する生物の中で最も古くから飛翔を行なっている生物の一つです。現存種だけでも世界で5000種以上が報告されており、さまざまな環境に進出し、さまざまな翅の形態を獲得しながら多様化しています。そのため、トンボ類の翅形態と生息環境や生態的特性との関係を調べれば、さまざまな飛翔特性を達成する翅形態が明らかになるかもしれません。もしかすると、工学的な応用にもつながるかもしれません(バイオミメティクス)。
その他のテーマ(過去のものも含む)
遺伝的多様性の空間勾配
種内多型のある生物では、型の比率がどの集団でも一定であることは少ないです。型の比率が空間的になだらかに変化する場合、そのような空間変異をクラインと呼びます。型の比率のクラインは多くの生物種で認められるものの、その成立の機構を実証した例はごくわずかです。なぜなら、型の比率のクラインの成立を証明するためには、各集団に多様性を維持する選択圧(平衡選択)と空間的に方向性選択の強さが異なることの両方を示さなければならないためです。私は、アオモンイトトンボとニワゼキショウの色彩多型を用いて、型比のクラインの成立機構を検証しました。
遺伝的多様性の維持機構
色彩多型はさまざまな分類群で認められる現象です。色彩はさまざまな選択圧を受ける可能性のある形質なので、野外で観察される色彩多型の多くは何らかの平衡選択によって維持されていると考えられています。しかし、それを実証した例は多くありません。私は、イトトンボや外来種のニワゼキショウを用いて多様性の維持機構を研究してきました。現在は、ショウジョウバエの行動変異に関する維持機構についても研究しています。
種内多型と表現型統合とその遺伝的基盤(進行中)
ある形質が変化すると、別の形質の最適値に影響することがあります。また、複数の形質が組み合わさることで、最適な振る舞いをできるようになることも少なくありません。形質はそれぞれが独立に変化するのではなく、互いに影響し合いながら進化するのです。このような複数の形質の統合的な進化は、表現型統合と呼ばれます。また、表現型統合を引き起こすような選択圧を相関選択と呼びます。最近では、トランスクリプトーム解析やゲノム解析をはじめとする分子生物学的手法を用いて、アオモンイトトンボの色彩多型に関わる遺伝子の探索を行なっています。このような解析を通じて、多型の進化の歴史や、上述の表現型統合の分子基盤を明らかにしたいと思っています。高橋迪彦氏が中心となり解析を進めています。